
Steve Hunter, ncsce.org
The vast majority of researchers seem to agree that true feathers are unlikely to have evolved more than once. (I believe that Evgeny Kurochkin is one of the few dissenters on that point.) Therefore, whoever finds the most likely precursor of feathers has also probably found the precursor of birds. We shall take a look at feathers, a couple of alternatives that have been proposed as possible "almost feathers" and at the way feathers have been treated by cladistics.
Before vetting proto-feathers, it is worthwhile to look at what a feather really is.
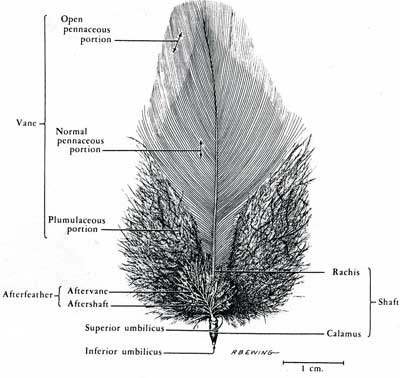
The vane is the "feather" part of the feather. Within the vane the shaft is called the rachis. The rachis is a robust, pith-filled shaft, which supports the barbs. The barbs, in turn, have a main shaft called a ramus, which supports the barbules.
The barbules of a cohesive, aerodynamic feather come in two types. The ones away from the base of the feather (distal) have little hooklets, which hook onto ridges on the barbules towards the base of the feather (proximal).
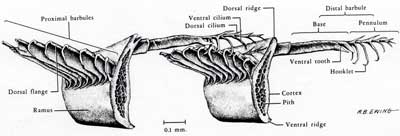
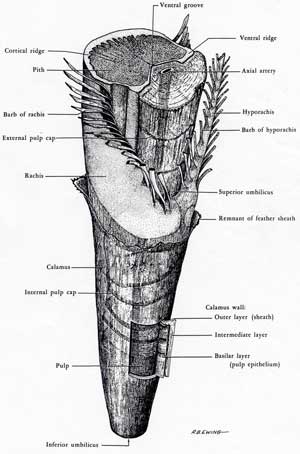
The base of the shaft of a feather is an hollow, cylindrical structure called a calamus. Inside the calamus, there are a series of hemispheric pulp caps, which are left over from the formation of the feather.
The afterfeather is an odd, miniature, mirror image of a feather growing out of its calamus. It is reduced to some stray barbs in many feathers and, even if it is well developed, it appears to be shed not long after the feather emerges from the sheath. I seem to be alone in suspecting that it may be evolutionarily significant. (Homologous to the posterior surface of the scale?) Time will tell on that one.
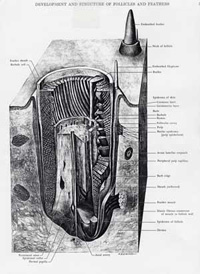
In extant birds, the follicle holds the feather erect. Additionally, it provides a unique interaction between the dermal and epidermal layers, which produces a series of feathers (often different sorts) as the bird grows and sheds its feathers in molting.
A mature feather is not living tissue. It is composed of structural protein in the beta keratin family. Alan Brush has made much of the uniqueness of the particular beta keratin and, on the basis of this uniqueness, has questioned the homology of reptilian scales and feathers.
In 2003, Roger Sawyer and colleagues publishedthe results of
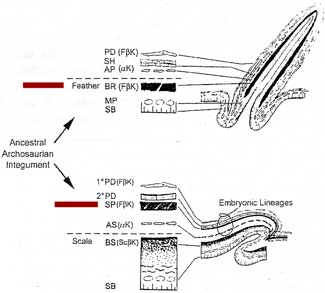
Feathers are highly derived scales. How evolution turned scales into feathers is the contentious question at hand. There are likely clues to this process in the way an individual feather grows, if we are clever enough to tease them loose.
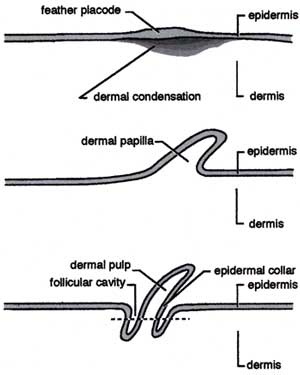
A feather begins as a cluster of elongate epidermal cells, the feather placode. The dermal condensation forms below. The basilar layer of the epidermis differentiates to add the intermediate layer of the epidermis, from which most parts of the feather are derived.
Initially, the dermal cells induce the differentiation of the epidermal cells and set growth rates and responses to hormones. Then the epidermis takes over. Apart from the ability to trigger feather growth, the dermis retains control over no other details of feather development.
An elongate feather bud emerges creating what has been referred to as a "finger of dermis covered by a thimble of epidermis." At this point it generally resembles a reptilian scale. In cross section, the thimble part resembles a tire tube of intermediate epidermal cells with thin layers of basilar and outer epidermis inside and out, respectively.
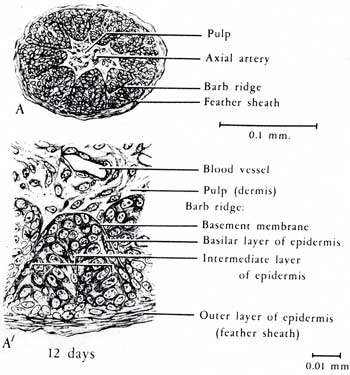
There is more rapid proliferation of the epidermal cells anteriorly, so the bud bends back. In the distal third of the bud, cells of the intermediate epidermis cluster on the interior surface of the dorsal side to form two adjacent longitudinal ridges. This is the first manifestation of the barb ridges.
There is further development of the follicle and the beginning of the formation of feather muscles. Beginning at the tip of the feather bud and progressing toward the base, the cells of the barb ridges begin to differentiate into cells that will become the ramus and barbules of the barbs and cells that will die off without keratinizing. The ridges that will become the rachis and hyporachis do not differentiate in this way. The core of the feather bud is a cylinder of pulp - dermal cells - with an axial artery that provides nourishment to growing feather.
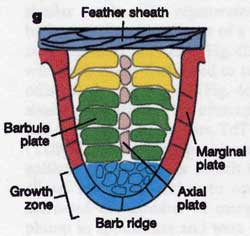
Once the follicle is fully formed, all new cells propagate from a "doughnut" of epidermal cells at the base of the follicle known as the epidermal collar. New cells are added at base of the incipient feather so that the cells that produce the calamus are added last.
Voila! A feather!
(This is obviously an extremely condensed description on of a reasonably complex process. For a more detailed treatment, please see Lucas and Stettenheim. And for more up to date research, please see the additional reading.)
"Feathered dinosaurs" took the world of vertebrate paleontology by storm in the late '90s and spilled out into the popular press in a series of breathless articles announcing the end of the debate over the origin of birds. In the cold light of day, some of these seem most likely to be birds and, therefore, that they are feathered is not particularly surprising. But some of these beasts do seem to be dinosaurs.
So, what of these feathers?

I had hoped to provide a better close-up photo of this stuff, but the best I could find is shown at the right. It appears to be filamentous. There have been tentative claims of some branching, but it is very difficult to distinguish a single structure from the jumble.
There have also been suggestions that the thick, dark, stubby structures along the bottom of the photo were something like calami. But there do not appear to be any finer filaments issuing from them. They appear to be stand-alone structures.


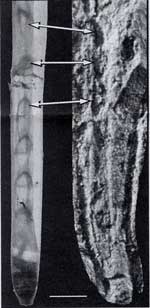
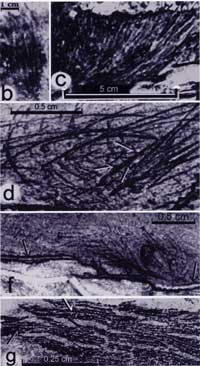
There have also been some good suggestions as to what these odd structures might have been if not proto-feathers.
John Ruben and his colleagues found that modern sea snakes and marine iguanas have bundles of collagen fibers that provide soft-tissue support for caudal fin-like structures. They are arrayed in a way that is remarkably reminiscent of the fuzz of Sinosauropteryx.
In 2003, Theagarten Lingham-Sollar published the results from his research into the preservation of collagen fibers of sharks and dolphins and on fossilized integumentary fibers in ichthyosaurs. Again he found among the variety of structures many characters that strongly resembled dino fuzz.
The skeleton is not in the best condition and is missing its pelvic girdle, hind limbs and tail. What makes it particularly interesting is the very unusual structures arrayed in pairs along the midline of its back. These structures also appear as isolated elements in the same formation.
For a few decades they were just called "elongate scales" and largely ignored.
Recently, Terry Jones and a number of colleagues have taken another look at these things. In a paper published in the 6/23/00 Science, they reported finding a number of remarkable, feather-like characters including an hollow calamus with pulp caps, a sheath and a generally feather-shaped vane supported by an apparently hollow rachis.
A follicle does not fossilize. But the combination all of the aforementioned characters strongly suggests growth from a feather-like follicle. (If this is true then these things on Longisquama fit Richard Prum's definition of a feather!)

I see most of the extraordinary, feather-like details (the calamus, the sheath, the rachis, etc.). But I can't make out barbs or barbules. What I see is a corrugated membrane.
Yu, et al., did do some work with the suppression of genetic and molecular signaling pathways to see the effects on feather development. When they suppressed sonic hedgehog, they found that the resulting feather had barb rami partially joined by membranes.
Paul Maderson has pointed out that one of the things that makes a feather unique is that the unfurled feather is substantially larger than the follicle that produces it. A scale is laminar. A feather is branched and larger than its follicle. Might the corrugated membrane of Longisquama represent something betwixt and between - a laminar structure that is larger than the follicle that produced it?
In contrast to the filamentous dino fuzz, there is an awful lot about the odd structures on Longisquama that does correlate with the details of feathers and feather development.


In their chapter in the book, Mesozoic: Birds Above the Heads of Dinosaurs, Clark and his colleagues present an extensive cladistic analysis of 44 species of dinosaurs and birds. Only one of the 208 characters used deals with feathers. By comparison, there are about a dozen dealing just with the ilium. Since the cladogram gives each character equal weight, the ilium, as an example, is twelve times more important than the feather in understanding the origin of birds.
I have reproduced the cladogram at the right with a color coding of the feather character. (Please click the image for a larger version that is easier to read.) Symmetrical feathers are coded as the primitive state (2 taxa), while asymmetrical feathers are coded as the derived state (2 taxa). All other taxa are coded as "?" meaning it is unknown whether or not they had any feathers. The authors do not include filaments in their data matrix.
As it sits, it seems pretty neat and straightforward. (Although it would mean that fully formed, but symmetrical, feathers are the primitive state for all therizinosaurs, oviraptorosaurs, troodonts and dromaeosaurs.)
But this does not represent the whole story as it is known at this point.
I accepted the filaments as the most primitive form of feather. They have been found on four taxa - Sinosauropteryx, Sinornithosaurus, Beipiaosaurus and Shuvuuia. I also included a second species of Microraptor that has been described by Xu, et al., as having asymmetric feathers. And I have included Longisquama.
This is not a re-running of the cladistic analysis, but an overlay of the feather data over the cladogram as it exists. The results are at the right. (Please click the image for a larger version that is easier to read.)
With the addition of just a little more data, the cladogram gets significantly stranger. Either there were lots and lots of reversals or pterosaurs, hadrosaurs, brontosaurs and even tyrannosaurs had the sort of partial feather found on Longisquama. And every case of dino fuzz represents a profound reversal. Even without Longisquama, three of the four instances of dino fuzz represent profound reversals.
Clark and his colleagues clearly would come up with different results if they had chosen to include all of the available feather data in their analysis and to give it proper relative weight.
The cladistics-only folks insist that their cladograms are the only objective means for understanding the history of life on the planet. But the results are contingent upon which characters and species are included and how they are coded and weighted. That can never be entirely objective.
Feathers are wonderfully complex structures that, as far as we know, are unique to birds. It seems to me that any analysis that seeks to find the origin of birds, but gives short shrift to feathers, is doomed to be half-assed.
(all are PDF files)
Chuong, Chodankar, Widelitz and Jiang, Current Opinion in Genetics & Development, 2000, 10:449-456
Chuong and Homberger, Journal of Experimental Zoology (MOL DEV EVOL) 298B:1-11 (2003)
Lingham-Soliar, Naturwissenschaften, 24 July 2003
Prum, Journal of Experimental Zoology (MOL DEV EVOL) 285:291-306 (1999)
Xu, Zhou, Wang, Kuang, Zhang and Du, Nature, VOL 421, 23 January 2003
Yu, Wu, Widelitz and Chuong, Nature, VOL 420, 21 January 2002
Chiappe and Witmer, University of California Press, 2002
Feduccia, Yale University Press, 1996
Jones, Ruben, Martin, Kurochkin, Feduccia, Maderson, Hillenius, Geist and Alifanov, Science, 23 June 2000, VOL 288, 2202-2205
Lucas and Stettenheim, Agricultural Handbook 362, U. S. Department of Agriculture, Washington, DC, 1972
Padian and Chiappe, Scientific American, February 1998, 38-47
Sawyer, Salvatore, Potylicki, French, Glenn and Knapp, Journal of Experimental Zoology (MOL DEV EVOL) 295B:12-24 (2003)
Scotchmoor, Breithaupt, Springer and Fiorillo, American Geological Institute, 2002




